|
Abstract FUNCTIONAL SYSTEM OF ANTlbRAVITY AS MECHANISM OF SELF-REGULATION OF AGE development PROCESSES WHEN TAKING UP PHYSICAL EXERCISES V.M. Chenegin, doctor of medicine, lecturer Volgograd state academy of physical education Key words: mechanism of age development, graviaf fe re n tat ion, functional system, acceleration-retardation, differences, cumulative effect of sports training. The aim of this paper is to expound a hypothesis about the functional system of antigravity as a mechanism of self-adjustment in age development processes which realizes their genetic depen dance and adjusts the influence of environment factors. According to the proposed hypothesis we (had) tried to explain the peculiarities of influence which different kinds of sports exert when such processes as age development and age formation of sexual and individual differences are considered. The possible area of application is also planned.
|

ФУНКЦИОНАЛЬНАЯ СИСТЕМА АНТИГРАВИТАЦИИ КАК МЕХАНИЗМ САМОРЕГУЛЯЦИИ ОНТОГЕНЕТИЧЕСКИХ ПРОЦЕССОВ ПРИ ЗАНЯТИЯХ ФИЗИЧЕСКИМИ УПРАЖНЕНИЯМИ Кандидат медицинских наук,
доцент В.М.Ченегив
Ключевые слова: механизмы онтогенеза, акселерация-ретардация, гравиафферентация, функциональная система, половой диморфизм, кумулятивный эффект спортивной тренировки. Предлагается для обсуждения рабочая гипотеза о функциональной системе антигравитации (ФСА) как о врожденном механизме саморегуляции и согласования важнейших онтогенетических процессов, а одновременно и опосредования влияния на них средовых факторов, и в частности физических упражнений (ФУ). Наше предположение о существовании ФСА нашло подтверждение в монографии [4]. ФСА в условиях жизни на Земле необходима для постоянного преодоления и компенсации сил земного тяготения, гравитации. Ее рецепторный компонент составляют: тельца Пачини, проприоцепторы, висцеральные баро- и хемоцепторы, вестибулярный аппарат [11, 25, 26, 27]. На основе гравицепторной информации в ЦНС формируются все компоненты ФСА, свойственные любой функциональной системе (ФС) по П.К. Анохину [1]: афферентный синтез, цель, акцептор и программа действия. В эфферентный аппарат ФСА входят: 1) скелетные мышцы, обеспечивающие позные реакции и антигравитационный компонент перемещений массы тела (МТ) человека; 2) системы циркуляции, дыхания, крови, тканевого метаболизма и выделения, участвующие в энергообеспечении функций эффекторов и в компенсации гидростатических сдвигов; 3) система организации поз и движений, а также трофических процессов во всех структурах организма [4, 15, 23, 25, 29]. Особого внимания заслуживают имеющиеся в цитированной литературе данные о воздействии гравиафферентации на трофические и пластические процессы в организме вообще и в опорно-двигательном аппарате, в частности. Именно эти процессы составляют основу ведущей для прогрессивной стадии онтогенеза закономерности - неуклонного увеличения размеров тела. Возраст и масса тела при менархе
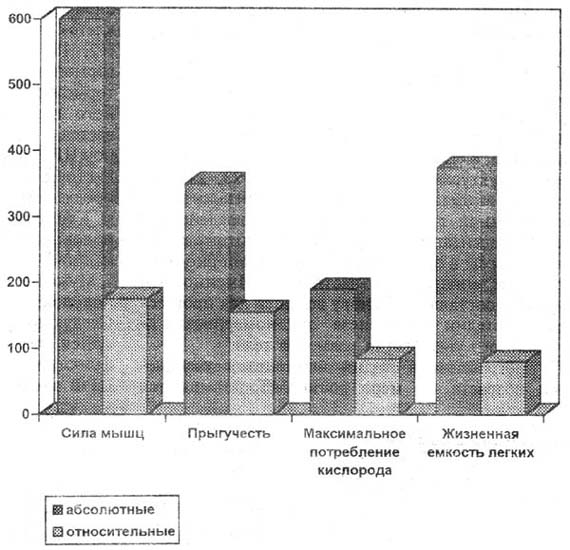
В прогрессивной стадии онтогенеза с увеличением возраста по мере роста МТ повьшается интенсивность гравиафферентации. Эффекторный аппарат ФСА соответственно усиливает стимуляцию трофических и пластических процессов в клетках, органах и физиологических системах, вовлекаемых в ФСА. Вследствие этого синхронно нарастают их размеры и функциональные возможности. И так продолжается по восходящей спирали из года в год до наступления генетически обусловленной биологической зрелости. Любые средовые факторы, в том числе и двигательная активность, вызывающие такие изменения роста МТ, которые достигают критической степени, сказываются на росте и формообразовании опорно-двигательного аппарата [6,10] и всего организма [24,28,30]. Это может осуществляться посредством отклонения динамики нарастания потока гравиафферентации от генетически программируемого и сказываться в вице акселерации или ретардации онтогенетических процессов. В литературе имеются некоторые данные относительно предложенной гипотезы. В биологическом созревании растущего организма ключевую роль играет [3,8,9,17] совокупность нервных клеток гипоталамуса, которую можно назвать центром репродуктивной функции (ЦРФ). У ребенка ЦРФ незрелый, и потому он обладает высокой чувствительностью к половым гормонам. В соответствии с законом об обратной взаимозависимости чувствительности и силы нервных клеток по отношению к процессам возбуждения [12, 18] даже очень низкая концентрация этих гормонов в крови вызывает у детей запредельное торможение ЦРФ. На начальном этапе пубертат-ного периода биологическое созревание сказывается в скачкообразном нарастании силы нейронов ЦРФ, проявляющемся в его растормаживании. Растормаживание ЦРФ запускает стимуляцию продукции гормонов по цепочке: гонад олиберины в гипоталамусе - лютропин в гипофизе - половые гормоны в гонадах (и, по-видимому, в надпочечниках). Мужские половые гормоны ускоряют не только половое развитие, но и общеростовые процессы. Следующий этап пубертатного периода характеризуется первичным включением репродуктивной функции (менархе, поллюции). Гипоталамический ЦРФ вновь затормаживается, продукция половых гормонов после небольшого снижения стабилизируется на высоком (дефинитивном) уровне. Окостеневают эпифи-зарные зоны роста, рост длины тела (ДТ) прекращается. В качестве фактора, обусловливающего наступление данной стадии полового созревания, называют достижение такого уровня физической зрелости, который обеспечивает полноценное осуществление функции воспроизведения потомства, а по некоторым из цитированных авторов [9] - достижение критического уровня МТ. Обусловленность включения репродуктивной функции достижением критического уровня МТ подтверждается наблюдениями о большей зависимости менархе от МТ, чем от паспортного возраста (см. таблицу). Приведенные материалы можно использовать как основание для предположения об обусловленности этапов созревания ЦРФ повышением силы его нервных процессов, повышением порогов возбудимости к содержанию половых гормонов, вызываемым нарастающим потоком гравиафферентации. Право на существование предложенной гипотезы подтверждает общеизвестная закономерность, состоящая в том, что в растущем организме максимальные проявления функциональных возможностей (МПК, МПС, ЖЕЛ и т.п.) увеличиваются преимущественно в абсолютном выражении, а относительные их величины (в пересчете на 1кг МТ) мало или совсем не изменяются. В качестве иллюстрации приводятся материалы лонгитудинальных исследований (рис. 1).
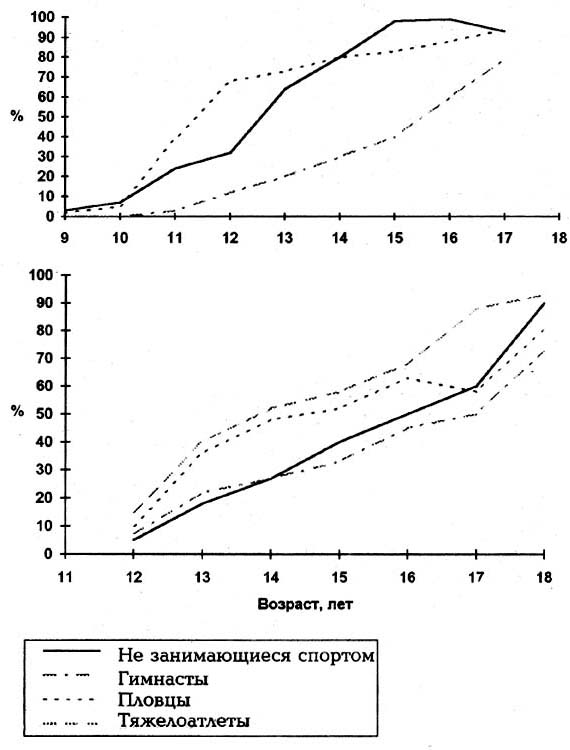
Рис. 1. Соотношение прироста абсолютных и относительных величин у девочек от 4 лет (100%) до возраста максимального развития признака 15 - 17 лет Доказательства предлагаемой гипотезы получены и в собственных исследованиях. У детей школьного возраста обоего пола, не занимающихся спортом, выявлена синхронность погодовых изменений скорости роста ряда антропометрических и функциональных показателей, наиболее заметная в пубертатном периоде, когда онтогенетические процессы ускоряются (рис. 2). Девочки:
Мальчики:
Рис. 2. Возрастная динамика скорости роста: I - % массы тела у девочек, не занимающихся спортом, и у занимающихся спортивной гимнастикой; II - % массы тела у мальчиков, не занимающихся спортом, и у занимающихся тяжелой атлетикой; III - % длины тела у девочек, не занимающихся спортом, и у гимнасток; IV - % длины тела у мальчиков, не занимающихся спортом, и у занимающихся тяжелой атлетикой; V - % жирового компонента у девочек, не занимающихся спортом, и у гимнасток; VI - % жирового компонента у мальчиков, не занимающихся спортом, и у занимающихся тяжелой атлетикой; VII - ЧСС в покое у девочек, не занимающихся спортом, и у гимнасток; VIII - ЧСС в покое у мальчиков, не занимающихся спортом, и у занимающихся тяжелой атлетикой Синхронность брадикардических сдвигов с пубертатным ускорением роста МТ отмечалась и другими исследованиями [12,19]. Самое большое снижение ЧСС наблюдается в первые 1-3 года [2,5], когда скорость роста МТ - наибольшая. Нейрофизиологической основой любого поведенческого акта, в том числе и произвольной мышечной деятельности, являются ФС. При занятиях спортом формируются Ф С специализированной спортивной подготовки (ССП). В ФС ССП как ФС высокого социального уровня включаются биологические ФС, в том числе и ФСА. Предполагается, что механизм ФСА является одним из ведущих в опосредовании влияния двигательной активности на онтогенетические процессы. Спорт высших достижений можно рассматривать как естественный эксперимент с применением во всех его видах нагрузок, в равной степени околопредельных, но со спецификой направленности в каждом из них. Для начала были выбраны два вида ССП: спортивная гимнастика и тяжелая атлетика. ФУ спортивной гимнастики можно назвать "телесными", поскольку мышечные усилия направлены на собственное тело (МТ): на удержание позы или перемещения тела. Повседневная ССП у юных гимнастов значительно усиливает поток гравиафферентации. Специальные тяжелоатлетические ФУ отличаются преобладанием направленности усилий на внешние предметы (штанга), их можно назвать "предметными"; они несравненно меньше усиливают гравиафферентацию. У гимнасток наблюдаются резко выраженная ретардация, значительная десинхронизация, а также сглаживание неравномерности течения онтогенетических процессов (рис. 2, 3). У юных тяжелоатлетов ретардировано только развитие жирового компонента и не происходит десинхро-низации онтогенетических процессов. Противоположная направленность воздействия двух видов ССП на онтогенетические процессы объяснима механизмом ФСА с учетом фазности влияния нарастающей интенсивности раздражителя [14,16]. Поскольку тяжелоатлетические ФУ мало усиливают гравиафферентацию, они не выводят ее за пределы оптимальных величин и составляют фактор стимуляции. Спортивная подготовка гимнасток ввиду ее преимущественно антигравитационной направлености настолько сильно увеличивает поток гравиафферентации, что она перехлестывает верхнюю границу оптимума, вызывает торможение гипоталамического ЦРФ. Саморегуляция онтогенетических процессов механизмами ФСА, несмотря на противоположную направленность тренировочных эффектов в рассматриваемых видах спорта, находится в пределах биологической целесообразности. Зависимость акселерирующего или ретарди-рующего влияния спортивной подготовки от той доли, которая приходится на антигравитационный компонент мышечной деятельности, носит, по-видимому, характер общей закономерности. Это отмечалось в материалах последующих исследований еще пяти групп: пловцов и прыгунов в воду ( мальчиков и девочек) и гимнасток-художниц. В плавании мышечные усилия направлены на преодоление гидродинамического сопротивления, они почти полностью "предметного" характера и поэтому мало усиливают поток гравиафферентации. Специального рассмотрения заслуживает половой диморфизм тренировочного эффекта. Вызванные спортивной подготовкой изменения темпов онтогенетических процессов, независимо от вида ССП, при их направленности в сторону акселерации, сильнее выражены у мальчиков, а ретардиру-ющий эффект тренировок преобладает у девочек (рис.3). Это проявление полового диморфизма вполне адекватно объясняется с позиции предложенной гипотезы, меньшей силой нервных процессов у женщин сравнительно с мужчинами. Выявленная в спортивных группах закономерность отмечается и у не занимающихся спортом в следующем виде: 1) более позднее начало пубертатного ускорения роста у мальчиков (с 12 лет) сравнительно с девочками (с 9 лет); 2) более позднее включение репродуктивной функции (поллюции у мальчиков - с 15 лет, а менархе у девочек - с 12,7 года) при более высокой МТ (мальчики - 60,3 кг, девочки - 47,7 кг); 3) течение ? ,^ пубертатного ускорения роста: у девочек в виде волн на 9-м, 11-м и 13-м годах, чередующихся с годами спада скорости роста, а у мальчиков в виде сплошного "скачка" с 12 до 15 лет (см. рис. 2). Все приведенные проявления полового диморфизма можно рассматривать как следствие более низкого у девочек и более высокого у мальчиков порога возбудимости гипоталамического ЦРФ к гравиафферентации.
Рис. 3. Динамика полового созревания (по уровню развития вторичных половых признаков, в % к дефинитивному, принятому за 100%) у девочек, не занимающихся спортом, и у гимнасток и пловчих. Динамика полового созревания (по уровню развития вторичных половых признаков) у мальчиков, не занимающихся спортом, у гимнастов, пловцов и тяжелоатлетов Приведенные материалы и доводы представляются достаточными, чтобы ставить вопрос о существовании ФСА как механизма саморегуляции онтогенетических процессов в прогрессивной стадии онтогенеза. В качестве заключения приводятся основные положения предлагаемой гипотезы. 1. Возрастное развитие МТ, сопровождающееся нарастанием интенсивности гравиафферентации, является ведущим онтогенетическим процессом, на котором конвергируются генетические и средовые обусловленности и который посредством ФСА направляет и согласует процессы биологического созревания растущего организма человека. 2. Исходя из роли ФСА в регуляции общеростовых процессов, в качестве главного критерия классификации ФУ, используемых в физическом воспитании детей, подростков и юношей, предлагается соотношение "телесной" (антигравитационной) и "предметной" направленности мышечных усилий, которым характеризуется интенсивность дополнительной (генерируемой ФУ) гравиафферентации. 3. Возрастные, половые и индивидуальные особенности физического состояния человека в пубертатных периодах (препубертат, пубертат и постпубертат) в значительной степени предопределяются различиями силы по отношению к процессам возбуждения гипоталамического центра репродуктивной функции. Механизмом ФСА осуществляется реализация генетической программы этих различий и корригирующих воздействий внешней среды. 4. С позиции гипотезы о роли ФСА в онтогенезе могут более адекватно решаться такие вопросы физического воспитания и спорта, как физкуль-турно-спортивная ориентация, определение возрастных рубежей сенситивных периодов и этапов спортивной подготовки, индивидуализация двигательных режимов и, по-видимому, многие другие. Литература 1. Анохин П.К. Биология и нейрофизиология условного рефлекса. - М.: Медицина, 1968. - 548 с. 2. Аршавский И.А. В кн.: Возрастная физиология. - Л.: Наука, 1975, с.5. 3. Баранникова И.А. В кн.: Общий курс физиологии человека и животных, кн.2. - М.: "Высшая школа". М., 1991, с. 72. 4. Белкания Г.С. Проблемы космической биологии, т.45. М., 1982. - 288 с. 5. Бирюкович А.А. В кн.: Функциональные и адаптационные возможности детей и подростков. т.1. М., 1974, с.70. 6. Буланова И.В. В кн.: Тр. Горьковского мед. ин-та, вып. 69, ч.1, 1977, с. 35. 7. Дильман В.В. Физиология человека, 1975, 1, № 2, с. 352. 8. Држевецкая И.А. Эндокринная система растущего организма. - М.: Высшая школа, 1987. - 207 с. 9. Дум игру И., Мэйкэнеску-Джоржеску М., Ротару М. и др. Физиология и патофизиология воспроизводства человека. - Бухарест: Мед. изд-во., 1981, 846 с. 10. Ковешников В.Г., Голод Б.В. Тр. Горьковского мед. ин-та, вып. 69, ч.1., 1977, с. 55. 11. Константинов А.И. В кн.: Общий курс физиологии человека и животных. - М.: Высшая школа, кн. 1, с.373. 12. Небылицин В.Д. Основные свойства нервной системы человека. - М.: Просвещение, 1966. - 383 с. 13. Преснякова А.М. Новые исследования по возрастной физиологии. 1984, № 1, с. 34. 14. Силла Р.В. Сб. докл. 2-го Респ. съезда эпидем., микробиол., инфекц., гигиенистов. Таллинн, 1972, с.150. 15. Ступаков Г.Н. и др. Космическая биология и авиакосмическая медицина, 1984, 18, №2, с.33 16. Сухарев А.Г. Тез. конф. "Мед. аспекты и пути оптимизации физ. восп. детей". Харьков. 1979, с.105. 17. Тэннер Д. В кн. : Биология человека. - М.: Мир, 1979, с. 412. 18. Теплов Б.М. Избр. тр. т.11. - М.: Педагогика, 1985. - 360 с. 19. Хрущев С.В. Врачебный контроль за физ. восп. школьников. - М.: Медицина, 1977, 288 с. 20. Ченегин В.М. Тез. докл. Всесоюзн. конф. "Современная морфология - физической каультуре и спорту". Л., 1987, с.153. 21. Ямпольская Ю.А. Автореф канд. дис. М., 1971.- 24 с. 22. Янкаускас И., Логвинов Э. Моторика растущего женского организма. - Вильнюс: Москлас, 1984. - 152 с. 23. Bonde-Petersen, Flemmmg-Acta astronaut, 1981, 8, N 9-10, 951 24. Hensgley L.D., East W.B., Stillwell У. L-Exercise and Sport, 1982, 53, N 2, 133. 25. Kozlovskaya J.B., Kreidich Ju.V., Organov V.S., Koserenko O.P.-Akta astronaut, 1981, 8, N9, 1059. 26. Metral S. Akta astronaut, 1981, 8, N 9-10, 1973. 27. Ross M.D. Aviat Space and Environ Med., 1984, 55, N 5, 16. 28. Rusconi R. Acta med. anvit, 1988, 20, N 2-3, 141. 29. Saiki H., Nakaya М., Sudon М., Jaketomi J. at. al. Acta astronaut, 1981, N 9-10, p. 959. 30. Williams Y.P.G. Growth and Develop. Pattern, 31, SuppL, 1981, p. 89. Поступила в редакцию 20.06.97
При любом использовании данного материала ссылка на журнал обязательна!
Реклама:
|
.jpg)
.jpg)