|
Abstract CRITICAL NOTES ON PROBLEM OF BIOPOWER SPECTRUM OF MODES OF RUNNING LOADING A.P. Kizko, Ph. D., associate professor E.A. Kizko, postgraduate Novosibirsk state technical university, Novosibirsk Key words: full biopower spectrum, criteria of substantiation of theory, method of estimation of "internal perfection" of theory. From the point of view of the modern poly system model of the substantiation of the scientific knowledge the theory divides into the levels based on the empirical and theoretical grounds. The usage of the theoretical criteria (and specifically the method of value of the "internal perfection" of the theory) is one of the most effective ways of checking the reliability of the theoretical knowledge and practical recommendations, which are based on it.
|

КРИТИЧЕСКИЕ ЗАМЕТКИ ПО ПРОБЛЕМЕ БИОЭНЕРГЕТИЧЕСКОГО СПЕКТРА РЕЖИМОВ БЕГОВОЙ НАГРУЗКИ Кандидат
педагогических наук, доцент А. П. Кизько Новосибирский государственный технический университет, Новосибирск Ключевые слова: полный биоэнергетический спектр, критерии обоснованности теории, метод оценки "внутреннего совершенства" теории. В 1994 г. была опубликована монография М. Р. Смирнова "Закономерности биоэнергетического обеспечения циклической нагрузки" [4]. В ней автор на теоретическом и эмпирическом материале обосновал "пути создания единой теории циклической нагрузки" на основе полного биоэнергетического спектра режимов беговой нагрузки. В 1996 г. вышло его учебное пособие "Теоретические основы беговой нагрузки" [5], рекомендованное научным сотрудникам, преподавателям и студентам институтов физической культуры и факультетов физического воспитания, а также тренерам-практикам. В рецензии на эту книгу отмечается: "В результате проведенного исследования автору удалось научно обосновать ряд новых закономерностей как биоэнергетического, так и спортивно -педагогического характера, определить конкретные пути развития и совершенствования предложенной им системы, а также пути реализации этой системы в реальной спортивной практике" [5, с. 2]. Обоснованность теории можно критиковать исходя из двух критериев. Первый критерий общепринятый: теория не должна противоречить данным опыта. То есть здесь речь идет о проверке теоретической основы на имеющемся или получаемом опытном материале. Как просто на первый взгляд выглядит это требование, но насколько тонким оказывается его применение. С одной стороны, "часто, если не всегда, можно сохранить данную теоретическую основу, если только приспособлять ее к факторам при помощи более или менее искусственных дополнительных предложений" [8, с. 526]. С другой стороны, объективность экспериментальных данных во многом зависит от правильной постановки задач опыта и методов ее решения. Второй критерий не имеет прямого отношения к опытному материалу, а касается предпосылок самой теории (основных понятий и основных соотношений между ними). Известно, что в качестве фундамента теория нуждается в некоторых общих положениях, отражающих определенные общие черты множества экспериментально установленных фактов. Согласно этим положениям в дальнейшем исследователь начинает развитие следствий. Как утверждают ряд авторитетных специалистов в области методологии теоретических исследований, анализ используемых в теории понятий и "выявление обстоятельств, от которых зависит их обоснованность, и того, как они возникают из данных опыта, не является праздной забавой" [7, с. 217], а служит одним из наиболее эффективных методов проверки правильности теории. Мы применили этот метод к оценке "внутреннего совершенства" теории циклической нагрузки М. Р. Смирнова. В результате анализа предпосылок, на которых строится вывод "фундаментальных закономерностей биоэнергетики", являющихся методической основой "научно обоснованных параметров практической подготовки спортсменов и физкультурников" [4, 5], выявлено две ошибки, которые существенно влияют на теоретическое содержание модели полного биоэнергетического спектра беговой нагрузки, предложенного автором, и соответственно на практические рекомендации. Рассмотрим первую из них, обнаруженную в области теоретических предпосылок автора при выводе закономерностей биоэнергетики человека. Для этого проследим логическую цепь переходов, сделанную М. Р. Смирновым, от первоначальных положений к теоретической модели исследуемого процесса. Из области биохимии автором было принято положение, согласно которому биоэнергетика мышечной деятельности исходит из того, что существует основной химический реагент, обеспечивающий функции миофибриллярной системы, - аденозинтрифосфорная кислота (АТФ), и есть несколько путей ее ресинтеза в организме человека [1, 9 и др.]. В этой связи М. Р. Смирнов формирует одно из основных положений теоретической концепции полного биоэнергетического спектра: "Все биоэнергетические процессы, характеризующиеся либо различными исходными субстратами, либо отличающимися условиями ресинтеза АТФ, могут рассматриваться как независимые источники энергии в организме человека" [5, с. 38]. Неоспоримым фактом является то, что большинство метаболических процессов в живом организме протекают параллельно и при этом взаимодействуют и дополняют друг друга. Для того чтобы "попытаться хоть как-то систематизировать взаимопереплетающиеся биохимические процессы и применить в теоретических расчетах метод математического моделирования" [5, с. 39], автор применяет понятие "принцип суперпозиции". Это позволило ему получить первую теоретическую предпосылку, которая гласит: "Существует ряд отдельных метаболических источников"[4, с. 44]. Из области теории химической кинетики и горения было принято положение, что "работа какого-либо источника энергии, сопровождающаяся расходом топлива или химического субстрата, изображается интегралом мощности по времени действия данного источника" [5, с. 39], т. е.
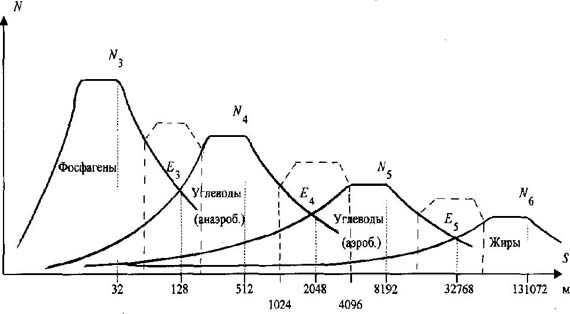
Это положение является второй теоретической предпосылкой. На основании теоретических предпосылок М. Р. Смирнов констатирует, что "существует принципиальная схема спектра метаболических источников, охватывающая весь диапазон мышечной деятельности" [5, с. 40]. Графическую интерпретацию спектра метаболических источников автор предлагает рассматривать с учётом принципа суперпозиции, в виде ряда "независимых кривых с частично взаимопересекающимися подинтегральными площадями" [5, с. 40] (рис. 1). Представленная на рис. 1 принципиальная схема биоэнергетического обеспечения циклической нагрузки, выраженная через динамику мощности метаболических источников, в дальнейшем рассматривается М. Р. Смирновым как теоретическая модель процесса реализации биоэнергетических ресурсов организма человека. Неадекватность этой теоретической модели реальному процессу метаболического обеспечения циклической нагрузки раскрывают следующие факты. Автор некорректно интерпретирует содержание принципа суперпозиции применительно к одновременно протекающим в организме человека метаболическим процессам. Общепринято принцип суперпозиции понимать как "результирующий эффект от нескольких независимых воздействий" или представляю щий собой "сумму эффектов, вызванных каждым воздействием в отдельности" [6, с. 1302]. В то время как М. Р. Смирнов считает, что "суть "принципа суперпозиции "" в данном случае сводится к тому, что он постулирует преимущественный вклад соответствующего источника в конкретный отрезок времени при работе конкретной мощности, но при параллельном течении других метаболических процессов" [4, с. 43]. Короче говоря, по мнению М. Р. Смирнова, принцип суперпозиции - это "преимущественный вклад", а не "результирующий эффект". В этой связи графическая интерпретация процесса одновременного протекания метаболических процессов, выраженных через динамику мощности этих источников, в соответствии с общепринятым пониманием содержания принципа суперпозиции будет
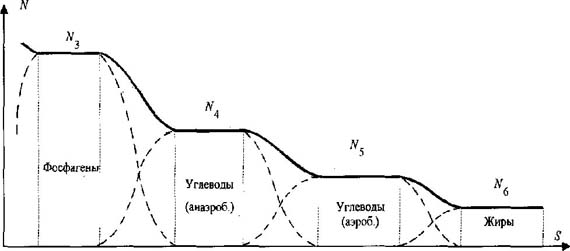
Рис. 1. Принципиальная схема метаболическо го обеспечения основного диапазона беговой нагрузки (сплошная линия - режимы, определяемые основными метаболическими источниками; пунктир - переходные режимы, определяемые дополнитель ными, промежуточными, источниками) отличаться от теоретической модели М. Р. Смирнова (рис. 2). Анализ "ряда независимых кривых с частично взаимопересекающимися подинтегральными площадями" [5, с. 40] с использованием понятий интегрального исчисления также фиксирует эту ошибку. Отметим ещё одно следствие этой ошибки. Из принципиальной схемы обеспечения циклической нагрузки (см. рис. 1) видно, что в переходном режиме между двумя основными энергоисточниками существует "провал" мощности. Как следствие этого, например при беге на 100 м, такие "провалы" мощности находили бы своё отражение в больших "провалах" скорости бега. Для коррекции теоретической модели М. Р. Смирнов был вынужден ввести в зонах метаболических переходов "промежуточные, переходные источники" [4, с. 59]. Однако реальная картина динамики суммарной мощности отдельных источников (см. рис. 2), построенная исходя из общепринятого понимания принципа суперпозиции и подкреплённая эмпирическим материалом (будет рассмотрено ниже), дает основание утверждать, что "промежуточные, переходные" источники не имеют права на своё существование. Зафиксируем результаты анализа теоретических предпосылок рассматриваемой теории. Во-первых, принципиальная схема биоэнергетического обеспечения беговой нагрузки М. Р. Смирнова, выраженная через динамику мощности отдельных метаболических источников (см. рис. 1), не отражает реального процесса (см. рис. 2). Во-вторых, количество отдельных метаболических источников уменьшается за счёт несуществующих "промежуточных, переходных" источников. Рассмотрим вторую ошибку, обнаруженную в области экспериментальных предпосылок вывода закономерностей биоэнергетики.
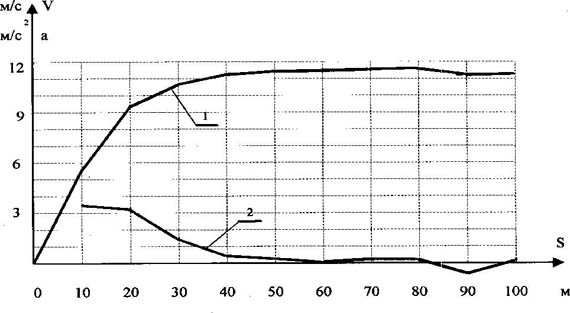
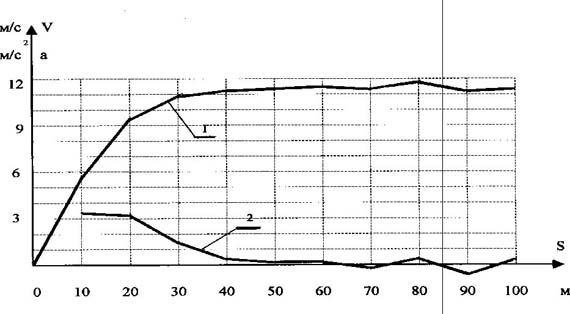
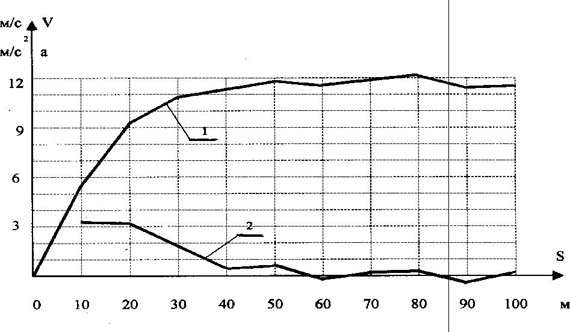
Рис. 2. Общепринятая картина понимания суперпозиционного взаимодействия в приложении к спектру биоэнергетических источников Известно, что все природные процессы протекают во времени и пространстве. В этой связи М.Р. Смирновым была поставлена задача согласовать характерные точки спектра метаболических источников (N3, E3; N4, E4; N5, E5; N6) с временными или метрическими параметрами перемещения спортсменов-мужчин, бегунов уровня международного класса. Точка N "ограничивает зону поддержания максимальной мощности данного источника (после точки N начинается постепенное понижение мощности)" [5, с. 42]. Точка E "открывает зону преобладающего действия последующего (менее мощного) источника" [5, с. 42]. Первой характерной точкой начала спектра (см. рис. 1) является точка N3 фосфагенного режима, которую М. Р. Смирнов определяет в результате анализа спидограмм элитарных бегунов (рис. 3-5). Автор считает, что "в районе 30-35 м спринтерской дистанции происходит резкий излом градиента скорости. Объяснить такой "перелом" спидограммы можно только появлением отрицательного градиента мощности превалирующего источника после точки N3" [4, с. 54]. В дальнейшем исследовании метрический параметр 30-35 м, соответствующий характерной точке N3 фосфагенного источника, М. Р. Смирнов рассматривает как одну из главных расчётных точек метрических параметров полного спектра метаболического обеспечения беговой нагрузки. Мы утверждаем, что обоснование, опирающееся только на зрительное фиксирование "резкого излома градиента скорости", некорректно и в итоге приводит к следующей ошибке. Известно, что перемещение спортсмена по дистанции обеспечивается силой мышечного сокращения. В области физики известна закономерность F = m · a. (1) Учитывая что за время пробегания дистанции 100 м масса бегуна (m) практически остается прежней, можно утверждать, что изменение ускорения тела (a) спортсмена будет адекватно отражать изменение силы мышечного сокращения (F (t)), т. е. F (t) = к · a (t), (2) где к - коэффициент пропорциональности. При пробегании соревновательной дистанции 100 м спортсмен фактически реализует свои максимальные способности. В этой связи можно считать, что величина силы мышечного сокращения будет лимитирована лишь мощностью использующихся метаболических источников. Это позволяет говорить о том, что изменение ускорения тела спортсмена на дистанции будет адекватно отражать и суммарную мощность метаболических источников, т. е. N (t) = к · a (t) или N (t) = к · dV/dt (t). (3) По данным спидограмм, приведённым М.Р. Смирновым [4, с. 52], нами были построены графики изменения скорости бега на 100 м трёх призёров Чемпионата мира 1991 г. (см. рис. 3-5). Исходя из условия, что a = dV/dt, рассчитаны и построены графики изменения ускорения этих спортсменов на данной дистанции (см. рис. 3-5). Формула 3 даёт основание утверждать, что изображённые на рис. 3- 5 графики изменения ускорения спортсменов необходимо рассматривать как реальную динамику изменения суммарной мощности метаболических источников, обеспечивающих перемещение спортсменов по дистанции. Обратим внимание на следующие вытекающие из анализа спидограмм реальные факты. У спортсменов до 15-20-го м до конца дистанции наблюдается площадка удержания максимальной мощности, что соответствует режиму работы одного из энергоисточников (см. рис. 2). Рубеж с 20-го до 60-го м представляет переходный режим, где вклад в энергообеспечение мышечной деятельности обеспечивается за счёт суммарного эффекта двух энергоисточников. С 60-65-го м дистанции и до финиша наблюдается площадка удержания мощности за счёт следующего энергоисточника. Изображённые на рис. 3-5 реальные изменения результирующей мощности метаболических источников являются, с одной стороны, убедительным подтверждением правильности нашей теоретической модели процесса энергообеспечения беговой нагрузки (см. рис. 2), с другой стороны, реальные метрические параметры характерных границ метаболических источников не согласуются с тем содержанием, которым их наполнял М. Р. Смирнов. Так, одному из главных расчётных параметров принципиальной схемы метаболического обеспечения беговой нагрузки (см. рис. 1) - рубежу 30-35 м - приписывалось "ограничить зону поддержания максимальной мощности данного источника" [5, с. 42], в то время как в реальном процессе этот рубеж - середина переходного режима между двумя энергоисточниками. Второй основной расчётный рубеж - 60-70 м - по логике автора открывает зону преобладающего действия "дополнительного, переходного" источника, которого в действительности не существует. Убедительно обоснован только рубеж 126 м.
Рис. 3. Спидограмма бега на 100 м Д. Митчела (ЧМ-91, Токио, рез. - 9.91 с) (1 - изменения скорости бега, 2 - изменение ускорения)
Рис. 4. Спидограмма бега на 100 м Л. Баррела (ЧМ-91, Токио, рез. - 9.88 с) (1 - изменения скорости бега, 2 - изменение ускорения)
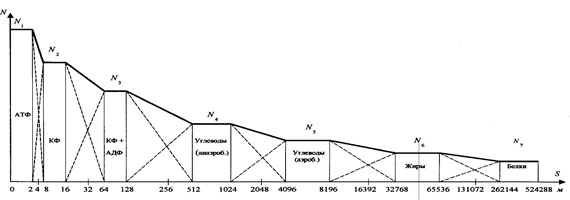
Рис. 5. Спидограмма бега на 100 м К. Льюиса (ЧМ-91, Токио, рез. - 9.86 с) (1 - изменения скорости бега, 2 - изменение ускорения) Тем не менее надо отдать должное М. Р. Смирнову, что даже на таких противоречивых экспериментальных предпосылках ему удалось построить достаточно убедительный "полный спектр метаболического обеспечения беговой нагрузки" и выявить фундаментальную закономерную связь между метрическими параметрами характерных границ метаболических источников в виде ряда членов геометрической прогрессии. Эта закономерность прослеживается и при анализе полученного нами реального процесса изменения результирующей мощности энергоисточников, если метрические параметры их характерных границ представить в виде таких значений: 16, 32, 64, 128. Эта последовательность значений представляет собой ряд геометрической прогрессии со знаменателем "2", что полностью соответствует открытому М. Р. Смирновым закону. Закономерная связь метрических параметров характерных границ метаболических источников и их мощностей (показано М. Р. Смирновым), выраженная числовым рядом членов геометрической прогрессии, имеет и теоретическое обоснование, которое получено нами при разработке универсального принципа развития и самоорганизации материальных систем [2, 3]. Экстраполяция закономерности метрических параметров характерных границ метаболических источников в левую и правую области от главных расчётных точек позволяет составить полное представление о количестве метаболических источников, способных обеспечить мышечную деятельность, от разового движения до непрерывной продолжительной работы. Количество метаболических источников в левой части спектра строго ограниченно и достаточно хорошо согласуется с циклами транспорта энергии в клетке (по В. Саксу и др.). В правой области ограничивающим этот ряд энергоисточником в настоящее время принято считать белки [1, 9 и др.]. Необходимо учитывать, что метрические параметры характерных границ энергоисточников рассчитаны на материале быстрейших людей планеты, которые в процессе спортивной тренировки подошли достаточно близко к максимально возможным границам развития систем, определяющим их результат. В этой связи метрические параметры характерных границ метаболических источников в правой области спектра нужно рассматривать как практически предельные значения их развития. На рис. 6 представлен полный спектр метаболического обеспечения беговой нагрузки, учитывающий ошибки, допущенные М. Р. Смирновым.
Рис. 6. Теоретическая модель полного спектра метаболического обеспечения беговой нагрузки Заключение. Обоснованность и соответственно значимость исследований в области теории и методики физического воспитания и спорта зачастую определяются традиционными стереотипами (но действительно объективными ) в форме нормативно-оценочных требований. В настоящее время эта область знания представлена совокупностью концепций, которые содержат в своем арсенале лишь экспериментальные критерии. Поэтому при сравнительной оценке теорий предпочтение отдаётся той, которая имеет большее эмпирическое содержание. В то же время современная методология познания отмечает ошибочность такого рода обоснования. Применение теоретических критериев, в частности метода оценки "внутреннего совершенства" теории, является одним из эффективных средств проверки достоверности теоретического знания и соответственно базирующихся на нём практических рекомендаций. Литература 1. Каменский М.И., Рогозкин В.А. Биохимия мышечной деятельности. - Киев: Здоровье, 1989. - 144 с. 2. Кизько А.П., Кизько П.А. Универсальный принцип развития и самоорганизации материальных систем: Учеб. пос. - Новосибирск: НГТУ, 1998. - 45 с. 3. Кизько А.П., Кизько П.А., Цимбалюк В.А. Методология анализа циклически волновых процессов развития: Учеб. пос. - Новосибирск: изд-во НГТУ, 2001. - 46 с. 4. Смирнов М.Р. Закономерности биологического обеспечения циклической нагрузки (на примере лёгкой атлетики). - Новосибирск: НГПУ, 1994. - 220 с. 5. Смирнов М.Р. Теоретические основы беговой нагрузки: Учеб. пос. - Новосибирск: НГПУ, 1996. - 217 с. 6. Советский энциклопедический словарь / Гл. ред. А. М. Прохоров. - 4-е изд., испр. и доп. - М.: Сов. энциклопедия, 1989. - 1632 с. 7. Эйнштейн А. Эрнст Мах: Сб. науч. трудов. Т. 4. Статьи, рецензии, письма, эволюция физики. - М.: Наука, 1967, с. 217. 8. Эйнштейн А. Автобиографические записки: Сб. науч. трудов. Т. 4. Статьи, рецензии, письма, эволюция физики. - М.: Наука, 1967, с. 520-531. 9. Яковлев Н.Н. Биохимия спорта. - М.: ФиС, 1974. - 288 с.
При любом использовании данного материала ссылка на журнал обязательна!
Реклама:
|