|
Abstract BIOENERGETICS OF RUNNING IN ENERGETIC COST UNITS E. A. Lazareva Ulyanovsk state university Key words: bioenergetics of muscular activity, running, time limit, power, energetic cost units The purpose of this paper was to describe the bioenergetics of running in the terms of energetic cost. The elite athletes (sprinters and stayers) took part in the experiment. The aerobic power output doesn't contribute to the general power production in running with tlim<27 s. The alactate and glycolic power production achieves its maximum level with tlim = 40 s (distance 300 m) and then it reduces. The speed of the power production decreases as tlim increases. When tlim = 9.5 min (distance 3000 m) the intensity of the energy output in anaerobic process is 1/50, but the speed of the energy output in aerobic process slowly increases with the increase of tlim (when tlim = 9.5 min achieves maximum in 280 cal/rg·min). On long distances of running the amount of energy, produced as a result of anaerobic processes is low, because of that these distances have aerobic trend by nature. If the increase of power of exercises takes place on long distances , it will be provided already by anaerobic mechanisms.
|

БИОЭНЕРГЕТИКА ЛЕГКОАТЛЕТИЧЕСКОГО БЕГА В ПОКАЗАТЕЛЯХ ЭНЕРГЕТИЧЕСКОЙ СТОИМОСТИ Э. А. Лазарева
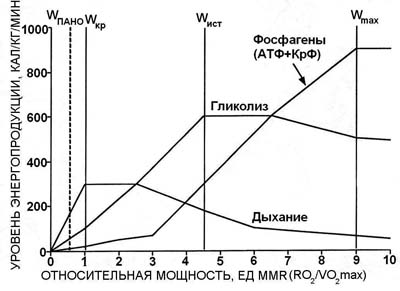
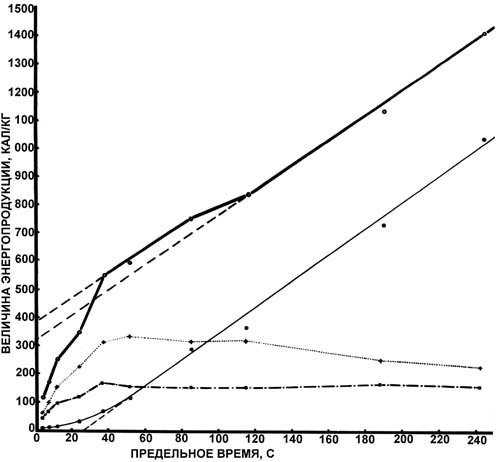
Введение. Успешное построение программы тренировочного процесса легкоатлета зависит от точности выявленных особенностей биоэнергетики отдельного спортсмена и адекватности применяемых к данному спортсмену методов тренировки. Любой из существующих в настоящее время видов тренировки определяет лишь общее направление тренировочного процесса, в то время как подготовка отдельного легкоатлета - это не простая реализация того или иного вида тренировки, а его адаптация к особенностям энергетической системы конкретного спортсмена. В связи с этим изучение особенностей биоэнергетики отдельного спортсмена - это ключ к пониманию логики построения тренировочного процесса, единственная возможность качественно изменить и усовершенствовать биоэнергетическую систему. Биоэнергетика легкоатлета - продукт сложного взаимодействия генетических и приобретённых качеств, знание особенностей которых (нормы реакции признака) может позволить установить оптимальный режим для совершенствования любого из тренируемых качеств. Иными словами, установление нормы реакции для любого признака должно предшествовать какому-либо воздействию на этот признак, показатель. Одним из наиболее обобщённых показателей биоэнергетики мышечной деятельности является количество выделенной в ходе упражнения энергии. Целью настоящей работы стало описание биоэнергетики легкоатлетического бега в терминах энергетической стоимости упражнения, а именно калорической стоимости бега на различные дистанции. Методы и организация исследования . В экспериментах принимали участие легкоатлеты высокой квалификации, специализирующиеся на преодолении коротких, средних и длинных дистанций. Эксперименты включали в себя выполнение упражнений (на беговой дорожке и велоэргометре) различной предельной продолжительности - 10, 15, 20, 30, 45, 60, 90, 120, 180, 360 и 600 с. С целью определения индивидуальных значений мощности выполняемой нагрузки при заданной предельной продолжительности использовали графоаналитический метод построения логарифмической зависимости "мощность-предельное время", исходя из результатов тестов максимальной анаэробной мощности [11] и удержания критической мощности [14]. Определение газового состава и объёмов выдыхаемого воздуха при проведении экспериментов на велоэргометре осуществляли при помощи мониторной системы ММС фирмы "Беккман" (США). Измерялись: уровень лёгочной вентиляции, уровень потребления кислорода, уровень выделения углекислого газа и значения дыхательного коэффициента. Экспериментальные данные обрабатывались компьютерной программой с целью получения значений О2-прихода, О2-долга и О2-запроса. Концентрация лактата в крови определялась калорическим [7] и энзиматическим [12] методами. Для установления значений биоэнергетических критериев мощности и ёмкости аэробных и анаэробных процессов испытуемы х тестирова ли на велоэргометре . Его программа включала следующие тесты, выполнение которых обеспечивало комплексную оценку аэробной и анаэробной работоспособности: 1) тест ступенчато возрастающей нагрузки, направленный на установление максимальной аэробной мощности, аэробной эффективности и суммарной анаэробной ёмкости [13, 9]; 2) тест удержания критической мощности, предназначенный для определения максимальных значений аэробной ёмкости [4]; 3) тест однократной предельной работы, ориентированный на определение мощности анаэробного гликолиза [2, 6]; 4) тест повторной предельной работы, исходя из которого оценивали гликолитическую ёмкость [2, 1]; 5) тест максимальной анаэробной мощности, необходимый для оценки максимальной алактатной анаэробной мощности [10, 8]. Результаты и их обсуждение. Первой и общепризнанной схемой, описывающей включение основных (фосфагенного, гликолитического, аэробного) биоэнергетических процессов в обеспечение мышечной деятельности, является схема, предложенная Н. И. Волковым [5]. Согласно данной схеме в пределах каждого из трёх биоэнергетических источников вырабатывается энергия (происходит ресинтез АТФ) и её количество зависит от предельного времени упражнения (дистанции легкоатлетического бега). Помимо этого количество выделенной энергии зависит от мощности (интенсивности) выполняемого упражнения. Иными словами, количество выделенной энергии лимитируется не только временем выполнения упражнения, но и скоростью преодоления дистанции. Взаимосвязь между уровнем энергопродукции в каждом биоэнергетическом источнике и относительной мощностью упражнения (в единицах MMR - RO2/VO2max) показаны на рис. 1. Согласно критерию мощности используемые физические нагрузки разделяются на нагрузки преимущественно аэробного воздействия, интенсивность которых не превышает значений порога анаэробного обмена; нагрузки смешанного аэробно-анаэробного воздействия {они разделяются на нагрузки субкритической интенсивности (уровень энергетических затрат не превышает значений максимального потребления кислорода) и нагрузки надкритической интенсивности, превышающие значения критической мощности}; нагрузки анаэробного гликолитического воздействия, интенсивность которых приближается к значению мощности истощения; нагрузки анаэробного алактатного воздействия с интенсивностью , близкой к значениям максимальной анаэробной мощности. Как показано Н.И. Волковым и И.А. Савелевым [3], энергетическое обеспечение мышечной активности до достижения значений критической мощности (соответствующей максимальному потреблению кислорода) осуществляется в основном за счёт аэробного окисления. В области напряжённой мышечной деятельности, где интенсивность упражнений превышает значения критической мощности, любое изменение мощности упражнения "оплачивается" исключительно усилением анаэробных процессов. Для анализа энергетики мышечной деятельности легкоатлетов нами взято достаточно большое количество разнообразных (эргометрических, биоэнергетических, биохимических) показателей. Максимально удобным было свести воедино данные о динамике этих показателей и представить их в качестве показателей, отражающих реальные энергетические затраты организма на бег на ту или иную дистанцию. Рассчитывалась как общая, так и специальная (алактатная, гликолитическая, аэробная) калорическая продукция. Расчёт алактатной калорической продукции производился исходя из зарегистрированных в эксперименте значений алактатного кислородного долга. С учетом того что ресинтез АТФ и КрФ при оплате алактатного долга осуществляется за счёт окисления углеводов и их производных, потребление кислорода на 1 моль ресинтезированных макроэргических соединений составит 3,53 л. В физиологических условиях калорический эквивалент 1 мл алактатного кислородного долга равен 2,82 кал. Таким образом, калорическая продукция алактатного анаэробного процесса вычисляется путём умножения величины алактатного кислородного долга на коэффициент 2,82. Калорическая продукция гликолитического анаэробного процесса рассчитывалась по данным замеров концентрации лактата на различных дистанциях легкоатлетического бега. Калорическим эквивалентом 1 г лактата является значение 222 кал. Для установления калорической продукции аэробного окисления использовали калорический эквивалент в виде значения кислородного прихода, равный 5, т.е. на 1 мл потребляемого кислорода выделяется 5 кал энергии (калорическая продукция аэробного процесса равна произведению кислородного прихода и коэффициента 5). На рис. 2 представлена зависимость общей и удельной энергопродукции от предельного времени (tпр.) бега. Кривая общей энергопродукции разделяется на три отрезка, которые могут быть описаны уравнением: E = a + b·tпр., где коэффициент a отражает размеры анаэробной энергопродукции, кал/кг, а коэффициент b - критический уровень энергопродукции, поддерживаемый за счёт аэробного источника энергии, кал/кг·мин.
Рис. 1. Взаимосвязь между уровнем энергопродук ции основных биоэнергетических источников и относительной мощностью упражнения
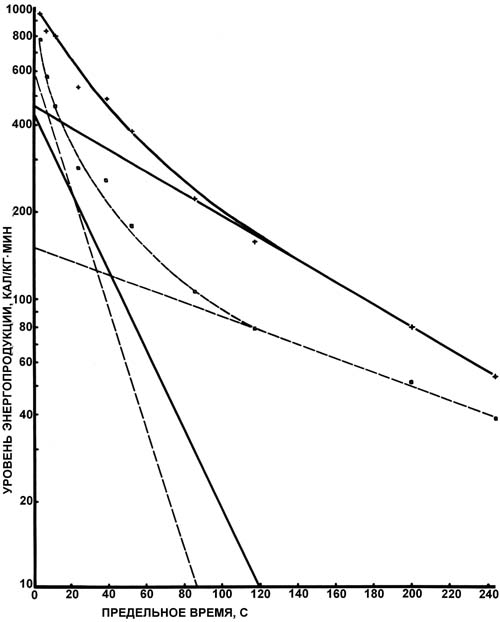
Рис. 2. График зависимости общей и удельной энергопродукции от предельного времени легкоатлети ческого бега. Условные обозначения: светлые кружки - значения общей энергопродукции, тёмные кружки - значения аэробной энергопродукции, крестики - значения гликолитической энергопродукции, квадратики - значения алактатной анаэробной энергопродукции На коротких дистанциях легкоатлетического бега линия общих энергетических затрат начинается в нулевой точке и характеризуется наклоном в 880 кал/кг·мин. Как можно видеть на графике, точки, соответствующие короткому спринту, находятся несколько в стороне от прямой линии - данный вид беговой нагрузки имеет непропорционально высокую энергетическую стоимость. Для дистанций легкоатлетического бега больше 800 м коэффициент b = 280 кал/кг·мин, что совпадает со средним значением максимума потребления кислорода нашими испытуемы ми. Наибольшее значение анаэробной энергопродукции (т. е. коэффициента a) на длинных дистанциях равнялось 320 кал/кг. Значение коэффициента b (критический уровень энергопродукции) на дистанциях от 300 до 800 м также составило 280 кал/кг·мин. Кривая аэробной энергопродукции (см. рис. 2), начиная с точки tпр.= 40 с (дистанция 300 м), идёт параллельно кривой общей энергопродукции на средних и длинных дистанциях легкоатлетического бега. На дистанциях легкоатлетического бега с tпр.< 27 с аэробная энергопродукция не вносит существенного вклада в общую энергопродукцию. Алактатная и гликолитическая энергопродукция (см. рис. 2) достигает максимума в точке с tпр.= 40-50 с и далее уменьшается. Наибольшая величина гликолитической энергопродукции в наших экспериментах составила 325 кал/кг, а наибольшая величина алактатной энергопродукции - 170 кал/кг. Скорость энергопродукции анаэробных процессов уменьшается по экспоненте по мере увеличения tпр.. В точке с tпр.= 9,5 мин (дистанция 3000 м) интенсивность выработки энергии в анаэробных процессах составляет 1/50 от такового значения в начальном периоде, в то время как скорость выработки энергии в аэробном процессе постепенно увеличивается с возрастанием tпр. (в точке tпр.= 9,5 мин достигает максимального значения в 280 кал/кг·мин). На полулогарифмическом графике, изображённом на рис. 3, можно видеть, что как алактатная, так и гликолитическая энергопродукция разбивается на два отдельных экспоненциальных участка. Первый экспоненциальный участок кривой алактатной энергопродукции характеризуется начальной скоростью 610 кал/кг·мин и временем полуспада (t1/2) 0,26 мин; ёмкость процесса - 228 кал/кг. Второй участок кривой характеризуется начальной скоростью 150 кал/кг·мин, временем полуспада 2,023 мин; ёмкость процесса - 437 кал/кг. Данные числовые значения позволяют заключить, что максимально возможная скорость алактатного анаэробного энергообразования выражается значением 760 кал/кг·мин, а максимальная энергетическая ёмкость - 665 кал/кг.
Рис. 3. Динамика изменений алактатного и гликолитического видов энергопродукции. Толстая линия соответствует уровню гликолитической энергопродукции, а тонкая - уровню алактатного анаэробного процесса Для первого экспоненциального участка кривой гликолитической энергопродукции начальная скорость составила 430 кал/кг·мин, время полуспада - 0,38 мин и энергоёмкость - 2,35 кал/кг. Второй экспоненциальный участок характеризуется начальной скоростью 466 кал/кг·мин, временем полуспада 1,26 мин и энергоёмкостью 847 кал/кг. В соответствии с полученными числовыми значениями максимальная мощность гликолитического процесса должна составить 896 кал/кг·мин, а общая энергетическая ёмкость - 1082 кал/кг. Это соответствует образованию 4,8 г лактата (эквивалентная концентрация в крови - 640 мг%). Концентрация лактата, рассчитанная по вышеприведённому графику, является "идеальной" концентрацией, которая могла бы быть достигнута . Если бы всё количество лактата оставалось неизменным , не происходило бы его утилизации мышцами и печенью. Полученные нами данные согласуются с результатами Н.И. Волкова и И.А. Савелева [3]. В кратковременных упражнениях высокой мощности уровень общих затрат энергии определяется кинетикой процессов анаэробного обмена. Скорость этих процессов достигает наибольших значений в упражнениях максимальной мощности, и она быстро снижается с увеличением предельного времени упражнения. Падение скорости анаэробной энергопродукции происходит неравномерно: оно резко выражено в первую минуту увеличения предельного времени упражнения (значение t1/2 для алактатного процесса составляет 18 с, а для гликолитического - 30 с) и менее заметно при больших интервалах времени работы. Скорость аэробного образования энергии непрерывно возрастает с увеличением предельного времени упражнения, приближаясь к своему наибольшему уровню - 1256 Дж/кг•мин при t1/2 = 180 с. В упражнениях максимальной относительной мощности, где tпр. не превышает 20 с, значение коэффициента Ep (ёмкость резервного источника энергии) равно нулю, а значение скорости высвобождения энергии в доминирующем энергетическом процессе (алактатном) составляет 9,0 кДж/кг•мин. В упражнениях субмаксимальной мощности, с tпр. от 20 до 180 с, ёмкость алактатного анаэробного процесса составляет 753 Дж/кг. Значение скорости высвобождения энергии в доминирующем метаболическом процессе (гликолизе) равно 2,26 кДж/кг•мин. В упражнениях большой мощности с tпр. от 3 до 10 мин ёмкость гликолитического анаэробного процесса равна 1,43 кДж/кг, а максимальная скорость высвобождения энергии в аэробном процессе - 1,25 кДж/кг•мин. Заключение. На дистанциях легкоатлетического бега с tпр.< 27с аэробная энергопродукция не вносит существенного вклада в общую энергопродукцию. Алактатная и гликолитическая энергопродукция достигает максимума при tпр.= 40 с (дистанция 300 м) и далее уменьшается. Скорость энергопродукции анаэробных процессов уменьшается по экспоненте по мере увеличения предельного времени. При tпр.= 9,5 мин (дистанция 3000 м) интенсивность выработки энергии в анаэробных процессах составляет 1/50 от такового значения в начальный период, в то время как скорость выработки энергии в аэробном процессе постепенно увеличивается с возрастанием tпр. (в точке tпр.= 9,5 мин достигает максимального значения в 280 кал/кг·мин). На длинных дистанциях легкоатлетического бега количество энергии, образованной за счёт анаэробных процессов, чрезвычайно мало, в связи с чем эти дистанции являются по своей природе работой чисто аэробной направленности. Если на длинных дистанциях будет иметь место увеличение мощности упражнения, это будет обеспечиваться уже анаэробными механизмами. Литература 1. Волков Н.И. Биохимический контроль в спорте: проблемы и перспективы // Теория и практика физ. культуры. 1975, № 11, с. 28. 2. Волков Н.И. Энергетический обмен и работоспособность человека в условиях напряжённой мышечной деятельности: Автореф. канд. дис. М.,1968. - 57 с. 3. Волков Н.И., Савелев И.А. Кислородный запрос и энергетическая стоимость напряжённой мышечной деятельности человека // Физиология человека. 2002, т. 28, № 4, с. 80 - 93. 4. Волков Н.И., Ширковец Е.А. Об энергетических критериях работоспособности спортсменов // Биоэнергетика. Л., 1973, с. 18. 5. Волков Н.И. Выносливость спринтера // Лёгкая атлетика. 1964, № 3, c. 28 - 31. 6. Волков Н.И. Тесты и критерии для оценки выносливости спортсменов. М., 1989. - 44 с. 7. Barker S.B., Summerson W.N. The colorimetric determination of lactic acid in biological material // J. Biol. Chem. - 1941. - Vol. 138. - P. 535. 8. Fox E.L. Measurement of the maximal alactic (phosphagen) capacity in man // Med. Sci. Sports. - 1973. - Vol. 5. - P. 66 - 71. 9. Hollman W. Hochstrund dauer leistungsfahigkeit des sportles. - Munchen: J. Bart, 1963. - 324 S. 10. Margaria R. Biomechanics and energetics of muscular exercise. - Oxford: Clarendon Press, 1976. - 184 p. 11. Margaria R., Aghemo P., Rovelli E. Measurement of muscular power (anaerobic) in mam // Eur. J. Appl. Physiol. - 1966. - Vol. 21. - P. 1662. 12. Methods of enzymatic analysis / Ed. Bergmeyer M.U. - N. Y.: Acad. Press, 1974. - 967 p. 13. Shephard R.J. Endurance fitness. Toronto: Toronto Press, 1977. - 714 p. 14. Volkov N.I., Shirkovets E.A., Borilkevich V.E. Assessment of aerobic and anaerobic capacity of athletes in treadmill running tests // Eur. J. Appl. Physiol. - 1975. - Vol. 34. - P. 121.
При любом использовании данного материала ссылка на журнал обязательна! |
 Ключевые слова: биоэнергетика
мышечной деятельности,
легкоатлетический бег, предельное
время, мощность, энергетическая
стоимость .
Ключевые слова: биоэнергетика
мышечной деятельности,
легкоатлетический бег, предельное
время, мощность, энергетическая
стоимость .